Immune regulation by the nervous system
It has long been proposed that various aspects of immune responses are influenced by nervous system activity. Indeed, lymphoid organs are innervated by various types of neurons, and immune cells express neurotransmitter receptors to respond to the neural inputs. However, little is known about how the inputs from the nervous system control immune responses. To solve this problem, we are studying the cellular and molecular basis for the neural regulation of immunity.
Adrenergic nerves constitute the efferent arc of the sympathetic nervous system and produce noradrenaline that induces cellular responses through α1-, α2-, β1-, β2- and β3-adrenergic receptors. Like other vital organs, lymphoid organs, including the bone marrow, thymus, spleen and lymph nodes, receive a rich supply of adrenergic nerves. We found that inputs from adrenergic nerves controlled lymphocyte egress from lymph nodes through β2-adrenergic receptors (J. Exp. Med. 2014). Activation of β2-adrenergic receptors expressed on lymphocytes enhanced the responsiveness of CCR7 and CXCR4, chemokine receptors that promote lymph node retention of lymphocytes, and consequently inhibited lymph node egress of the cells (Fig.1).
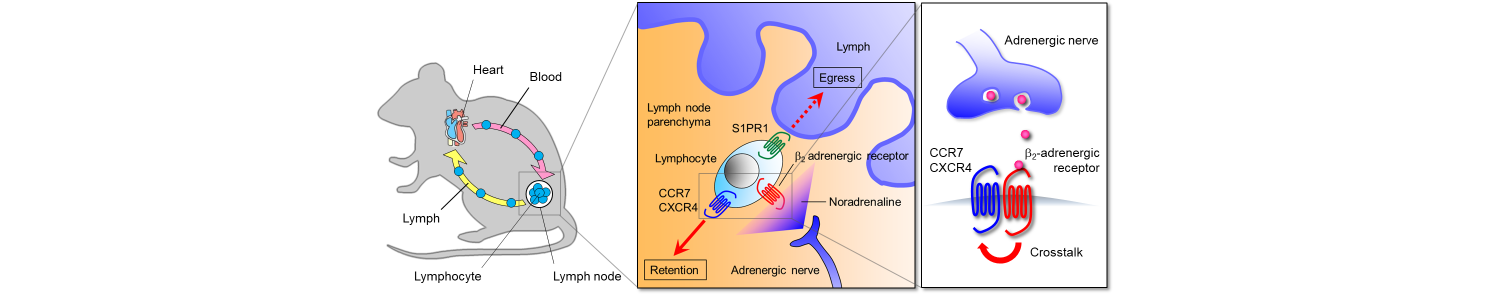
Figure 1. Adrenergic control of lymphocyte egress from lymph nodes
The activity of adrenergic nerves displays a circadian rhythm that is synchronized with the rest-activity cycle of the species. The noradrenaline release from adrenergic nerves increases during the daytime in humans, whereas it reaches a peak at night in rodents. We found that the night time surge of adrenergic nerve activity in mice restricted lymphocyte egress from lymph nodes, leading to an increase of lymphocyte numbers in lymph nodes. Immunization during the period of lymphocyte accumulation in lymph nodes enhanced antibody responses. The diurnal variation of the humoral immune response was dependent on adrenergic inputs and was diminished when lymphocyte recirculation through lymph nodes was stopped. These findings suggest that adrenergic control of lymphocyte trafficking contributes to daily fluctuations of adaptive immune responses (J. Exp. Med. 2016). Our study demonstrates that the immune system is poised to mount higher responses in lymph nodes during the active phase in a day, which may have evolved to maximize the efficiency of host defense when encounters with pathogens are more likely to occur (Fig. 2).
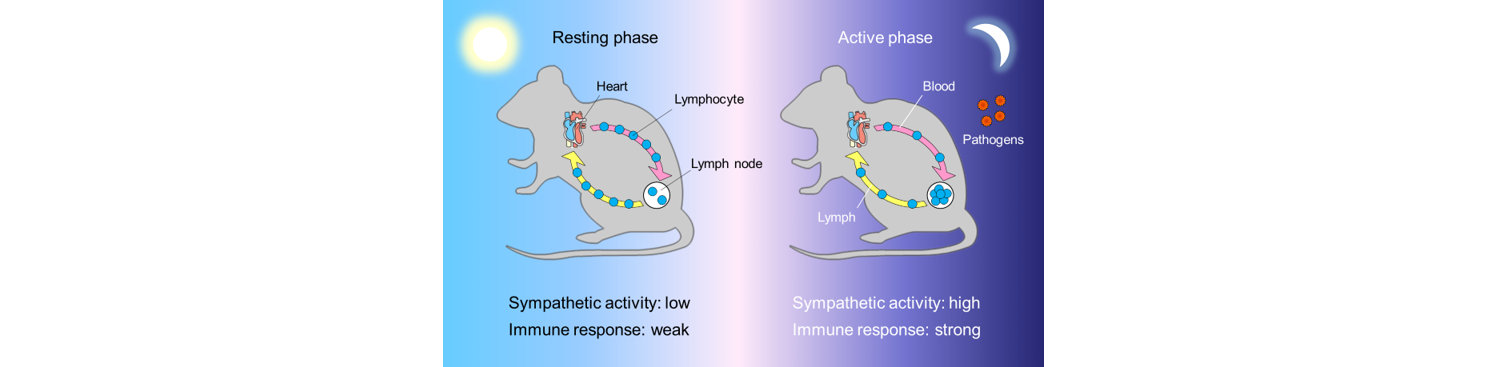
Figure 2. Diurnal control of adaptive immunity by adrenergic nerves
Targeting lymphocyte migration in autoimmunity
Lymphocyte migration is mediated by G protein-coupled receptors (GPCRs) that respond to chemoattractants, represented by chemokines. Agonist-bound GPCRs are phosphorylated by GPCR kinases (GRKs) and subsequently recruit β-arrestins that serve as scaffolds to activate signaling molecules, including mitogen-activated protein kinases (MAPKs). The GRK family consists of seven mammalian members, among which GRK2, GRK3, GRK5, and GRK6 are expressed ubiquitously. Different GRKs phosphorylate distinct sites on the C-terminal tail of the receptor, establishing a barcode that dictates the functional consequences of β-arrestin engagement. Thus, specific targeting of GRKs to activated GPCRs is crucial for signal transduction. However, the molecular mechanism that determines the specificity of GRK targeting is poorly understood.
In search of factors involved in GRK recruitment to chemoattractant receptors, we identified a protein complex consisting of copper metabolism MURR1 domain-containing (COMMD) 3 and COMMD8 (COMMD3/8 complex), which interacts with the C-terminal tail of chemoattractant receptors, including CXCR5, CCR7, and the oxysterol receptor EBI2, and promotes lymphocyte migration mediated by these receptors (J. Exp. Med. 2019). Mechanistic analysis for the action of the COMMD3/8 complex demonstrated that this protein complex functions as an adaptor that selectively recruits GRK6 to chemoattractant receptors in a GRK2/3-dependent manner, which promotes MAPK activation and consequently lymphocyte chemotaxis (Fig. 1). It has been suggested that the specificity of GRK recruitment to GPCRs is determined by the relative expression levels of individual GRKs, which vary among cell type, and distinct receptor conformations induced by ligand binding. Our study has identified a GRK-recruiting adaptor, the COMMD3/8 complex, as an additional determinant of GRK specificity for GPCRs.
Consistent with the positive regulatory role of the COMMD3/8 complex in lymphocyte chemotaxis, B cells deficient in this protein complex showed multiple defects in their migration in vivo. Additionally, deficiency of the COMMD3/8 complex severely impaired humoral immune responses. Therefore, the COMMD3/8 complex is essential for proper functioning of the immune system. Pharmacological inhibition of the COMMD3/8 complex may provide a novel approach for the treatment of immune disorders.
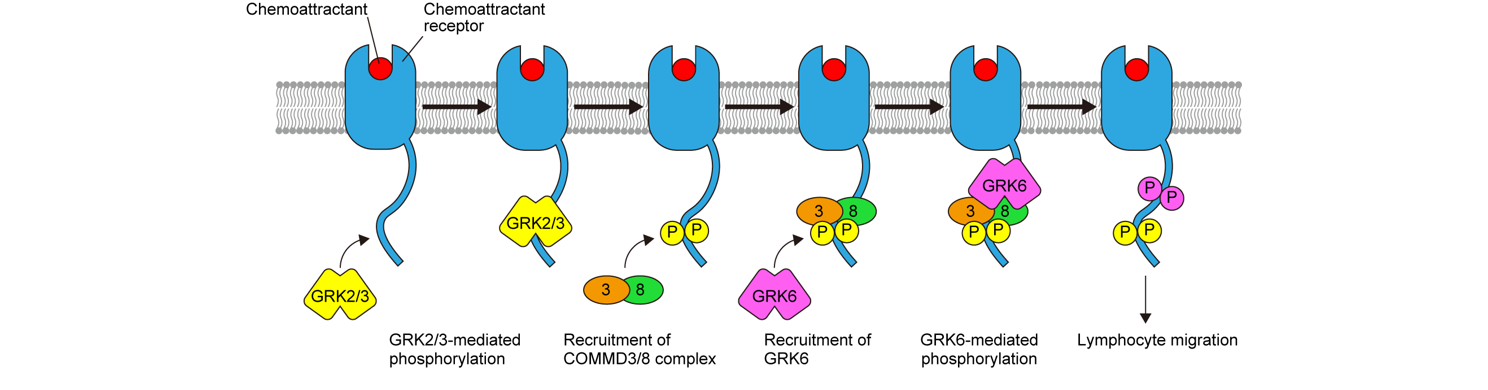
Figure 3. Role of the COMMD3/8 complex in chemoattractant receptor signaling
Based on the important role of the COMMD3/8 complex in humoral immune responses, we investigated the involvement of the COMMD3/8 complex in autoimmune diseases. Deficiency of the COMMD3/8 complex in collagen-induced arthritis, a B cell-dependent mouse model of rheumatoid arthritis, blocked disease progression, which was accompanied by reduced humoral immune responses to collagen. These observations suggest that the COMMD3/8 complex contributes to the pathogenesis of the autoimmune disease (Sci. Immunol. 2023).
We next searched for inhibitors of the COMMD3/8 complex that could be used for the treatment of autoimmune diseases. Screening of the chemical compounds that interfered with the association between COMMD3 and COMMD8 identified celastrol, an anti-inflammatory agent with a poorly defined mechanism of action. Celastrol suppresses B cell migration, humoral immune responses, and collagen-induced arthritis, recapitulating deficiency of the COMMD3/8 complex. These effects were abolished in mice expressing a celastrol-resistant mutant of the COMMD3/8 complex. These observations suggest that the COMMD3/8 complex is a major target of celastrol (Sci. Immunol. 2023).
Above findings establish the COMMD3/8 complex as a novel therapeutic target for autoimmune diseases and support consideration of celastrol as a lead pharmacologic candidate.
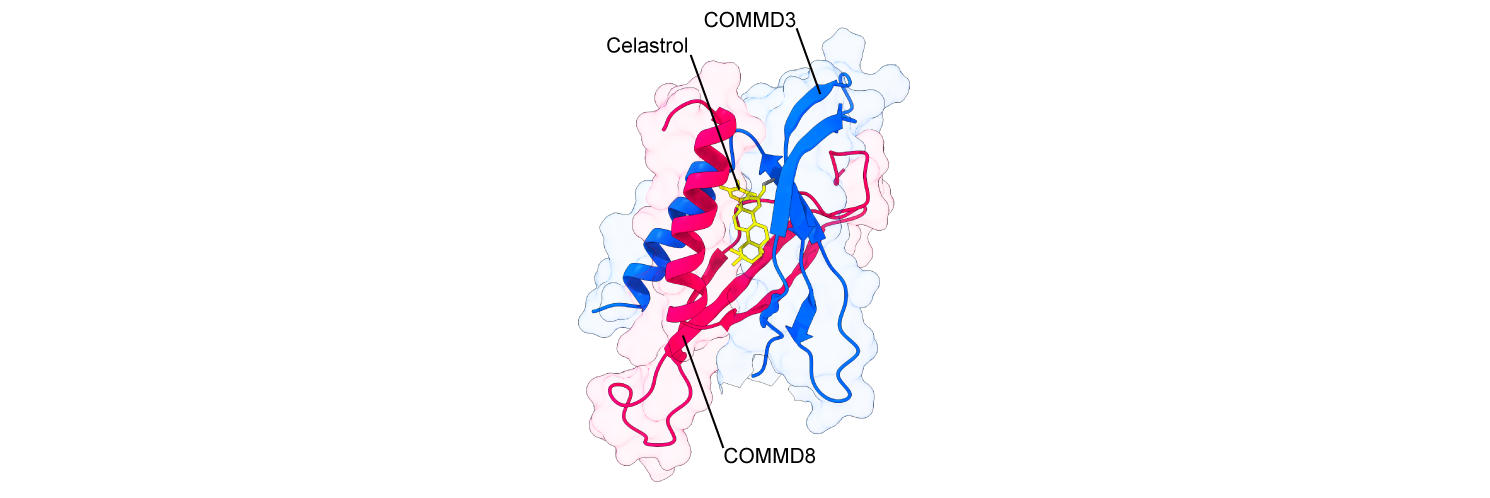
Figure 4. Model of the celastrol-bound COMMD3/8 complex
